Introduction
Scale: Bar=1mm unless noted. Click on images for larger version.
All photos by Bruce McCune unless noted.
Vegetative Structures
Habit
- Variations:
- erect
- erect to suberect
- appressed
- appressed to suberect
- appressed rarely pendulous
- appressed to pendulous
- pendulous
- erect or trailing
- erect or appressed

H. austerodes

H. wilfiana

H. apinnata

H. duplicata
Diameter
MaxSizeExt (cm): Extreme maximum diameter or length of the thallus (largest observed).
Branching
- Variations:
- isotomic dichotomous
- anisotomic dichotomous
- variable
- strongly pinnate

H. imshaugii

H. krogiae
Budding
- Variations:
- absent or rare
- occasional
- present

H. occidentalis - Budding adventitious branches

H. enteromorpha - Adventitious branches and budding

H. heterophylla - Lobes
Upper Surface Color of Thallus
Most of the characteristically brownish species (such as H. austerodes and H. bitteri) have gray to greenish gray shade forms. Other species (such as H. mollis) maintain a white to light gray color, even in extremely exposed habitats. Most of the species that are typically gray to greenish gray can be almost white to a relatively dark greenish gray, depending on the environment.
- Variations:
- white to greenish gray
- white
- gray
- gray to greenish gray
- greenish gray
- greenish gray to brown
- brown
- white to greenish gray or brown
- pale gray to black
- white to gray often dark mottled
- gray often dark mottled
- yellowish green

H. canadensis - Habit

H. rugosa - Lobes

H. inactiva - Upper surface solarized
Dark Mottles
- Variations
- none or rare
- sometimes present
- often present
- present

H. beringiana - Habit

H. fistulosa - Upper surface
Black Border
- Variations:
- not visible
- sometimes present
- often present
- present
- conspicuous

H. occidentalis - Lobes

H. oceanica - Lobes
Lobe Arrangement
- Variations:
- contiguous
- contiguous to +/- separate
- contiguous to imbricate
- separate
- contiguous or separate
- separate to centrally subcontiguous
- separate to centrally contiguous
- separate rarely contiguous
- separate to +/- imbricate


H. imshaugii - Habit

H. inactiva - Habit
Lobe Outline (or Profile)
- Variations:
- even
- even to +/- nodulose
- even to nodulose
- nodulose
- pinched and swollen
- no elongate lobes
- even to +/- pinched/swollen

H. apinnata - Habit
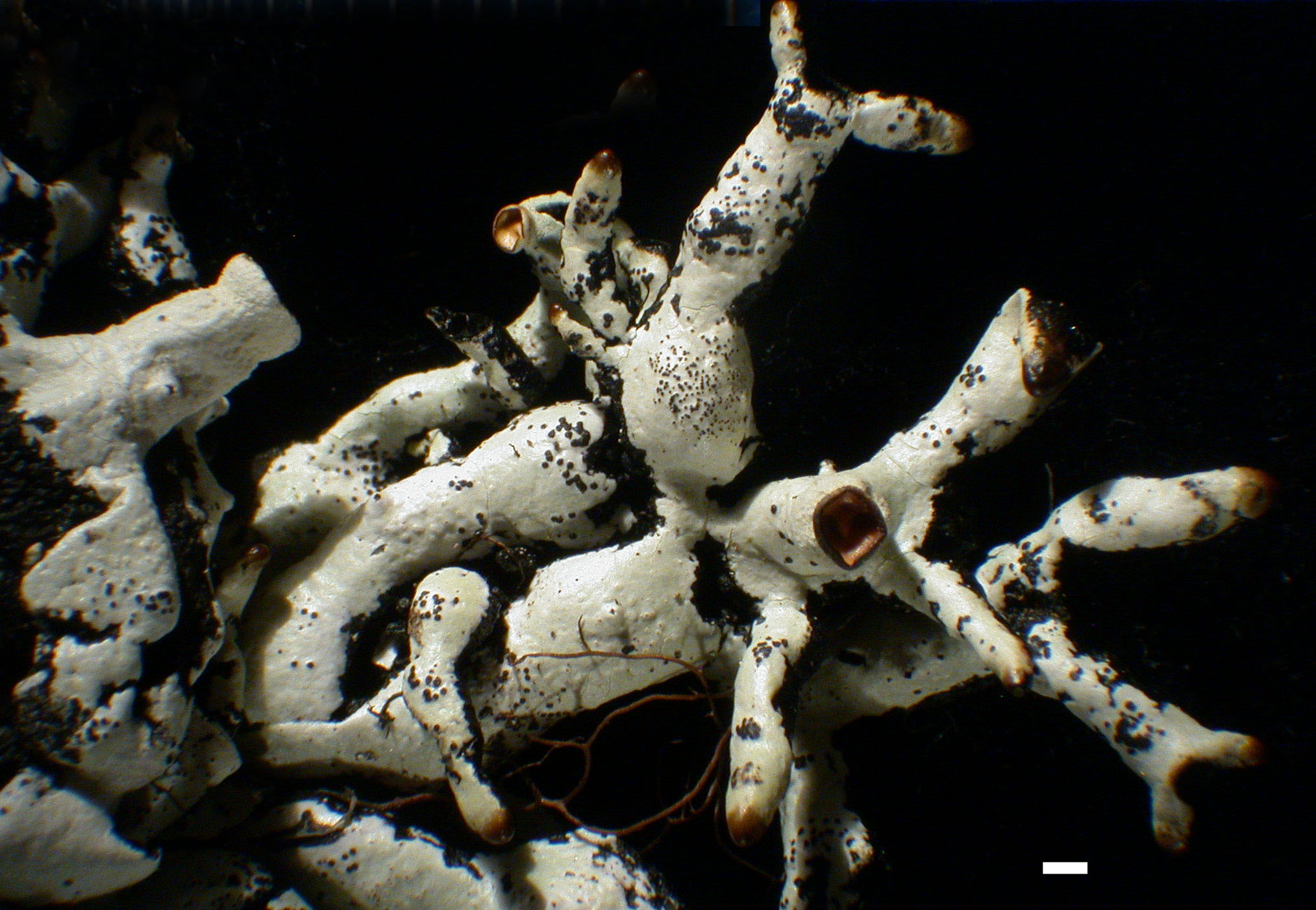
H. enteromorpha - Habit
Lobe Surface
- Variations:
- smooth
- smooth to weakly rugose
- occasionally rugose
- becoming rugose
- polygonal cracked
- becoming verrucose
- smooth with angular depressions

H. canadensis - Habit

H. occidentalis - Upper surface rugose
Lobe Pruina

H. lophyrea - Apothecia pruinose
Lobe Width Min-Max (Extreme)
- LobeMin: Lobe width (minimum) in mm.
- LobeMax: Lobe width (maximum) in mm. This value is a typical maximum size on a given specimen. Lobes are measured at a typical spot along the internodes, not at the often expanded portion at the branch points.
- LobeMaxExt: Lobe width (extreme maximum) in mm. The width of abnormally broad lobes is recorded here.
Lobe Width/Height Ratio Min-Max
- Min: (unitless) Minimum ratio of width to height of lobes, measured about 1 cm back from the lobe tip, but not at a node. To find this value select one or more lobes that are relatively thick but narrow. Make a transverse section either by cutting with a razor blade or tearing.
- W/H Max: (unitless) Minimum ratio of width to height of lobes, measured about 1 cm back from the lobe tip, but not at a node. To find this value select one or more lobes that are relatively broad but thin.
Lobe Tips Perforate
The perforations in Hypogymnia mostly follow one of two developmental patterns. In one the perforations are terminal on the lobe tips. When the lobe tip branches dichotomously, the perforation becomes axillary and the new apices elongate. Then the new apices can develop perforations. This pattern leads to many terminal and axillary holes, but only a few holes in the lower surface. In some species, such as H. heterophylla, the perforations are slow to develop, so that lobe tips mainly have pinholes, while large holes are seldom seen until the tip has become an axil. The fraction of lobes developing perforations is quite variable, but typically in the range of 5-75%. Example species with frequent tip perforations include H. apinnata, H. enteromorpha, H. occidentalis, H. sachalinensis, and H. vittata.
In the second developmental pattern the holes form on the lower surface just behind the apices (subterminal) or at the apices. As the lobe elongates, the hole is left on the lower surface. This results in a series of holes laid down on the lower surface, but with few truly terminal or axillary holes. Examples are mostly Asian species, including H. arcuata, H. hengduanensis, and H. yunnanensis. The holes may be so close as to commonly fuse, as in H. arcuata or the extreme case, H. fragillima, where the lower surface is typically more hole than tissue.
- Variations:
- perforate
- often perforate
- sometimes perforate
- sparsely perforate
- rarely perforate
- entire

H. heterophylla - Perforate lobe tips

H. occidentalis - Perforate lobe tips
Lower Surface Perforated
In most species the perforations are simple holes or with an inrolled edge. In a group of Asian species (e.g. H. macrospora), however, the rolled edge of a hole often forms a raised rim around the hole.
- Variations:
- heavily perforate
- perforate
- sparsely perforate
- sometimes perforate
- rarely perforate
- entire

H. inactiva - Perforate lobe tips

H. vittata - Lower surface
Cavernulae

H. hultenii - Lower surface with cavernulae

H. lophyrea - Lower surface with cavernulae
Thallus Texture
- Variations:
- cartilaginous
- papery
- soft
Medulla
- Variations:
- solid
- mostly solid
- partially solid
- rarely solid
- hollow

H. inactiva - Lobe cavity

H. tubulosa - Lobe cavity
Ceiling
- Variations:
- dark
- dark to grayish
- grayish
- grayish to white
- brownish to white
- white
- white or dark

H. apinnata - Lobe cavity

H. occidentalis - Lobe cavity
Floor Color
- Variations:
- dark
- dark to grayish
- grayish
- grayish to white
- white
- white or dark

H. farinacea - Lobe cavity

H. wilfiana - Lobe cavity
Reproductive Structures
Soredia
The presence and form of soredia are very important in differentiating among Hypogymnia species. The most distinctive type are produced in labriform soralia in burst lobe tips, as in H. physodes and H. vittata. In this case the soredia are produced initially on the medullary surface of the lobe cavity. Terminal but externally-produced and often capitate soralia are produced by H. tubulosa. Production of these soralia is often preceded by a darkening of the upper cortex at the lobe tip, sometimes with an oily sheen. In juvenile thalli these incipient soralia can be very helpful in distinguishing H. tubulosa from permanently esorediate species. Other kinds of soralia differ in developmental pattern and final appearance. I have designated 5 types of soralia, apart from those already described.
Type 1 soralia are terminal on short, often upturned, lateral lobes. These are frequent on H. bitteri. These lobes are typically narrower and shorter than the main lobes. The soralia are discrete, often smaller in diameter than the lobe on which they are borne (though occasionally broadening and becoming capitate). These soralia are produced by a gradual dissolution of the cortex, often preceded by a discoloration of the cortex.
Type 2 soralia are also terminal but are produced on the main lobes. They are corresponding larger than type 1 soralia. The soralia are typically less discrete than type 1 soralia and often occur on downturned lobes, like sorediate knuckles on the perimeter of the thallus. These occur in H. bitteri, rarely in H. austerodes, and frequently on the putative hybrids from H. bitteri and H. austerodes from the central and southern Rocky Mountains of North America.
Type 3 soralia are produced along edges of flakes of the upper cortex. These are characteristic of H. farinacea, H. pseudophysodes, and several other species. Frequently these flakes arise from polygonal cracks or rugosity. In the latter case, the cortex breaks along the microridges on the thallus, followed by splaying apart of the broken edges of cortex. The flakes do not penetrate all the way through to the hollow center of the medulla, but do appear to carry at least a thin layer of medullary hyphae.
Type 4 soralia develop by hemispherical semi-corticate warts on the upper surface. They blur the distinction between soredia and isidia, because the cortex appears to be at least partly intact. At fairly high power with a stereoscope, one can see the color of the cortex lightened by a subtle appearance of fine, pale markings. Over time these warts often becoming compound by repeated budding. This form of soralia is most frequently seen in H. austerodes. Occasionally the budding is so pronounced as to suggest true isidia.
Type 5 soralia develop by a gradual deterioration of the upper cortex, simultaneously removing the shiny luster of the upper cortex and becoming minutely roughened. Gradually these rough laminal areas develop soredia, usually in poorly soralia. This form of soralia is frequent in H. austerodes, but also occurs in many other species that produce laminal soredia.
- Variations:
- none
- present
- rare
- Type 1
- Type 2
- Type 3
- Type 4
- Type 5
- labriform
- terminal capitate
- laminal capitate

H. bitteri - Lobes

H. physodes - Soredia

H. physodes - Soredia

H. vittata - Apothecia

H. mollis - Soredia

H. tubulosa - Soredia

H. hultenii - Soredia

H. tubulosa - Soredia
Isidia
True isidia are rare in Hypogymnia. The best examples of true isidia in Hypogymnia are the cylindrical isidia found in the Asian species, H. duplicatoides, H. hengduanensis, and H. zeylanica. The isidiose soredia (type 4) found in H. austerodes approach globose isidia in form and probably in function. Species that typically produce small lobules, such as H. subobscura and H. kosciuskoensis, occasionally produce globose to clavate "isidia". However, given the preponderance of dorsiventral lobules in those species, rather than true isidia, perhaps they are best considered aberrant lobules.
- Variations:
- none
- present
- rare
- cylindrical
- globose
- clavate

H. austerodes - Isidia grading into soredia, note areas with roughened cortex

H. subobscura - Isidia
Lobules
- Variations:
- none
- rare
- often present
- present

H. krogiae - Lobules
Apothecia
Apothecia Frequency
- Variations:
- unknown
- rare
- occasional
- common
.JPG)
H. enteromorpha - Habit

H. occidentalis - Habit
- Variations:
- sessile
- substipitate
- stipitate
- sessile to substipitate
- substipitate to stipitate
- ApoDiamMax (mm): Diameter of the apothecial disk (maximum) in mm. This value is a typical maximum size based on examining numerous thalli.
- ApoDiamExtreme (mm): Diameter of the apothecial disk (extreme maximum) in mm. The width of abnormally broad apothecia is recorded here.
The lower surface of the receptacle (or stipe) often appears scabrous under the dissecting scope. Under the compound scope, the surface is covered with filamentous branched projecting hyphae, the filaments about 8-13 um wide and coated with POL+ crystalline material. The filaments form a mat typically about 50-65 um thick.
- Variations:
- urn shaped
- funnel shaped
- urn or funnel shaped
- n.a.
- Variations:
- absent
- sometimes
- present
- collapsed base
- loosely filled
- n.a.
- Variations:
- brown
- brown to reddish brown
- brown to dark brown
- reddish brown
- reddish brown to dark brown
- dark brown
- tan
- yellowish
- brown to olive
- pale olive to grayish
Spore Min-MaxWidth x Min-MaxLength
- SporeLengthMin (µm): Ascospore lengths and widths are measured outside of the asci, in water or 10% KOH, avoiding the runts (aberrant and immature spores). The minimum value is the low end of the typical range in size. Published spore sizes rarely mention the medium in which they were measured. I have found that visibility of spore characters, paraphyses, and hymenium often improve markedly after application of KOH. In Hypogymnia I have not observed a marked change in spore size with application of KOH.
- SporeLengthMax (µm): See above; the maximum value is the high end of the typical range in size.
- SporeWidthMin (µm): See above.
- SporeWidthMax (µm): See above.
Hypothecium
Immediately below the subhymenium is what we term the hypothecium. The upper part of the hypothecium is a compact tissue that can contain POL+ crystals(bright in polarized light). In other species these crystals are lacking, sparse, or concentrated in the base of the hypothecium. The distribution and abundance of POL+ crystals appears to be a useful species-level character in Hypogymnia. Below the hypothecium is the medullary part of the thalline exciple, which is always somewhat occluded by crystals and POL+ grayish white, and generally somewhat looser than the hypothecium.
Pycnidia
Pycnidia (frequency)
Frequency of production of pycnidia is highly variable in Hypogymnia, but can be fairly constant within species. Production of pycnidia appears to be strongly correlated with production of apothecia.
- Variations:
- unknown
- rare
- sparse
- occasional
- common

H. apinnata - Lobe tips

H. heterophylla - Perforate lobe tips
Schizidia

H. schizidiata - Schizidia propagules
Spermatia Shape
- Variations:
- rod
- rod to weakly bifusiform
- weakly bifusiform
- bifusiform
- SpermatiaLengthMin (µm): Spermatia (conidia) are measured in the same way as the ascospores, though we find them difficult to measure accurately under the microscope. We have therefore switched to measuring them with Photoshop from pictures at 400X. See details in McCune and Wang (2014).
- SpermatiaLengthMax (µm): See above.
- SpermatiaWidthMin (µm): See above.
- SpermatiaWidthMax (µm): See above.
Chemistry
Medullary spot tests

H. enteromorpha - P+orange-red

H. physodes - P+orange-red
The substance responsible for the K+ reddish brown reaction is 3-hydroxyphysodic acid. The reaction is rather curious and unlike the typical K reactions. This spot test appears to be quite reliable for detecting this substance. The reaction is first apparent wherever there are cracks in the cortex, initially (within about 30 seconds or less) turning the cracks brown, but then the cracks and upper surface gradually reddens (showing through the upper cortex). It takes about 30 minutes for the color to develop fully, but it rarely is necessary to wait more than a couple of minutes to be sure. So the color goes from an initial brown to a distinctly reddish brown.
The K+ red-brown reaction can also be seen on acetone extracts on a white spot plate, where the resulting color can be closer to orange. It can also be demonstrated on undeveloped spots on TLC plates. In the latter case, when the reagent is painted on with a soft brush, the 3-hydroxyphysodic acid spot turns a russet brown.
A weaker, slower, K+ orangish brown reaction is sometimes given by physodalic and protocetraric acids.
In rare instances in Hypogymnia (especially in the H. imshaugii group) one finds the medulla of certain species tinted by yellowish pigments, especially below the apothecia. In at least some cases these are K+ deep red-purple, suggesting the presence of anthraquinones.

H. canadensis - K+ reddish brown reaction

H. physodes - K+ reddish brown reaction

H. inactiva - KC+ red spot test

H. occidentalis - KC+ red spot test
Cortical Spot Tests
TLC
- Atranorin (and chloroatranorin)
- Physodic acid
- 3-hydroxyphysodic acid
- 2'-O-methylphysodic acid
- Physodalic acid
- Protocetraric acid (always with physodalic acid)
- Hypoprotocetraric acid
- Diffractaic acid
Ecological Characters
Substrate
- On Bark or Wood
- Host
For epiphytic species, list genera on which it grows. - Rock
- On Moss, Detritus or Sod